氮,是我们每天必摄入的营养物质,它以我们熟知的氨基酸形式存在于蛋白质之中,如我们餐桌上常见的鸡蛋、牛奶、豆奶、肉类等均含有丰富的氮。可是,这些蛋白质中的氮元素又是从何而来呢?
微生物造就循环之旅
在我们脚下的土壤中存活着许多我们用肉眼看不到的微小生命体,科学家们把它们称为微生物;在我们人类出现前,这些微生物已在地球上繁衍生息了数亿年,它们当中的大多数靠取食腐烂的动植物尸体存活,也有一些栖息在活的动植物体内。虽然,它们个体微小,但是数量却很惊人,小小的一捧土壤中就有上亿个微生物,因此,它们的生命活动不容小觑。土壤中的碳、氮、磷等化学元素的转化都是由它们来完成的,从这一点来讲,土壤微生物可以说是地球工程师。
氮是动植物生长的必需营养元素,它主要以气态形式存在于大气中,要经过三个过程,即生物固氮过程、硝化过程和反硝化过程,才能完成它的轮回,并为植物根系吸收利用,然后,再供给动物和人类;上述的三个过程均是由微生物完成的。

图1 氮循环示意图
第一段旅程:氮气分子变成氨基酸和蛋白质
大气中存储着丰富的气态氮分子,但遗憾的是这些氮分子是惰性的,植物和动物自身不能直接利用;只有在微生物与植物协作下,才能将这些惰性氮气转变为生物可利用的活性氮,进入植物体内,转变为氨基酸和蛋白质,如大豆和花生;参与这个过程的微生物叫固氮菌,这个过程也被称之为固氮作用。一般而言,固氮菌可分为三大类群,即自生固氮菌、共生固氮菌和内生固氮菌。
自生固氮菌是指那些在土壤中能够独立进行固氮作用的细菌。因为固氮菌的固氮酶对氧分子极为敏感,其活性的发挥必须在无氧条件下进行。自生固氮菌历经数亿万年,进化出了一套固氮酶的防氧保护机制,能够在进行有氧生活的同时固定空气中的氮。这些固氮菌保持很强的呼吸活性,借以维持细胞质内固氮酶周围的低氧环境;另外,它们的细胞中还含有一种铁硫蛋白,这种蛋白能与固氮酶结合,增加后者的稳定性。除此之外,自生固氮菌的细胞壁外层有一薄层荚膜或粘质,也能够阻碍氧气分子向细胞内扩散。
一些被称为根瘤菌的杆状细菌能与豆科植物共生,在豆科植物的细根中形成根瘤并固定空气中的氮气,供给豆科植物氮营养元素,因此营养学家们常常建议我们多食用豆类,满足人体对蛋白质的需求。这类根瘤菌能促使豆科植物根系异常增生,而且这些小家伙能以体表的鞭毛运动,主动寻找豆科植物,从豆科植物根毛侵入根内形成根瘤(图2),并在根瘤内转变为分枝的多态细胞,我们称之为类菌体。
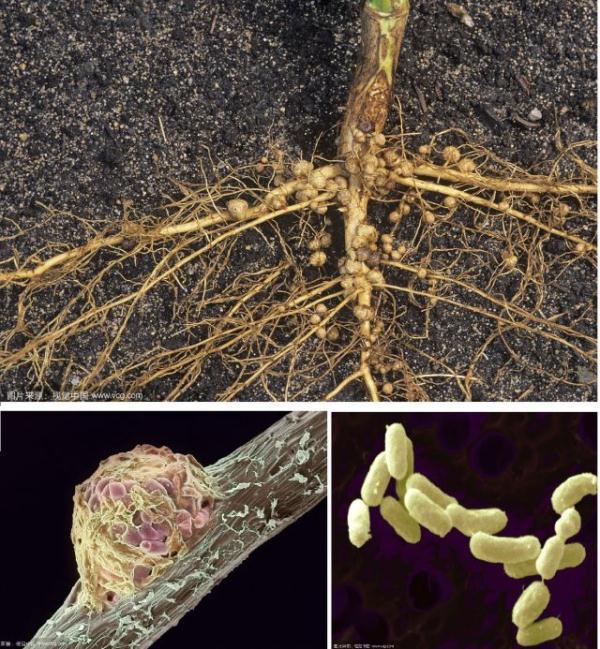
图2 根瘤与根瘤菌
另外一些非豆科植物体内也存在一些具有固氮能力的细菌,这些植物内生固氮菌通常能定殖在健康植物体内,与宿主植物进行联合固氮,这些多为土壤杆菌属,能够通过植物根系的伤口入侵多种双子叶植物和裸子植物,致使植物细胞转化为异常增生的肿瘤细胞,产生根癌、毛根或杆瘿等。
第二段旅程: 蛋白质蜕变为硝态氮
植物和动物死亡后,体内的蛋白质会被微生物降解为氨态氮,氨态氮经由硝化作用变为硝态氮。硝化作用可分为两个阶段:氨态氮氧化为亚硝态氮的氨氧化阶段,亚硝态氮氧化为硝态氮的亚硝酸盐氧化阶段;其中氨氧化作用,也被称为亚硝化作用, 是硝化作用的第一步, 同时也是氮循环的限速步骤, 对全球氮循环起决定作用。
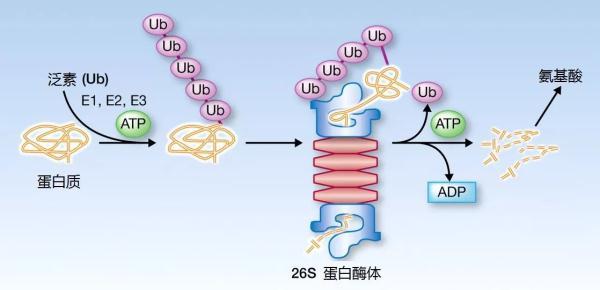
图3. 蛋白质降解
一百多年来,人们普遍认为土壤中的氨氧化主要是由一些化能自养型的变形菌—即氨氧化细菌催化完成的,主要包括亚硝化单胞菌、亚硝化球菌、亚硝化螺菌和亚硝酸叶菌等。
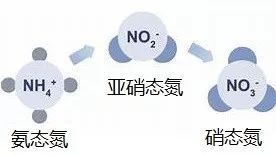
图4. 氨氧化
在土壤中, 目前所知的自养氨氧化细菌分别属于亚硝化单胞菌属和亚硝化球菌属, 是β-变形菌亚纲的一个单源进化簇,有别于海洋中的氨氧化细菌。自养型氨氧化细菌很难在实验室内被分离培养,科学家们只能借助于微生物分子生态学方法,通过解码16SrRNA基因或氨单加氧酶编码基因来推测其在生态系统中的分布和数量。
在2004年前后,对于氨氧化微生物认识发生了转变,科学家们利用一种新进发明的宏基因组学技术发现,海洋中生活的一些古菌的基因组中含有类似细菌编码氨单加氧酶的基因,而后2005年,科学家们又从美国西雅图的水族馆海水中分离培养到一株氨氧化古菌,彻底颠覆了人们对氨氧化微生物的认识。
2006年,来自挪威卑尔根大学的雷宁格博士和斯盖尔博教授在英国《自然》杂志上报道了它们的研究发现,土壤中具有氨氧化功能的古菌数量可达氨氧化细菌的3000倍。紧随其后,众多研究证实氨氧化古菌广泛分布于包括陆地、海洋和淡水湖泊中, 并且个体数量远高于氨氧化细菌。
第三段旅程:硝态氮又变为氮气
反硝化作用顾名思义是与硝化作用相反的氮转化过程,是硝态氮还原为气体氮素的过程。
反硝化作用将硝酸盐还原为气态产物, 是土壤中氮元素轮回的重要环节。这个过程非常复杂:NO3→NO2→NO→N2O→N2,从硝酸盐到氮气释放,需要4个连续反应步骤, 由反硝化细菌分泌的硝酸盐还原酶、亚硝酸盐还原酶、一氧化氮(NO)还原酶和氧化亚氮(N2O)还原酶四种酶类催化完成。
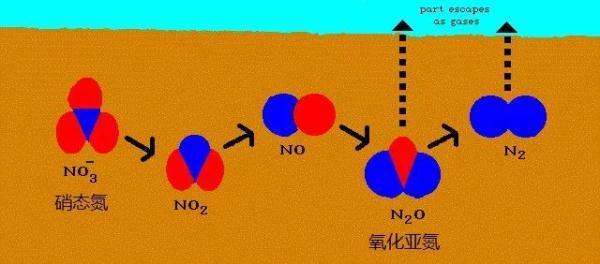
图5. 反硝化过程释放氧化亚氮气体和氮气
起初人们发现反硝化细菌只能存活于无氧的环境中,但是,最近发现许多有氧气环境中的微生物能在其细胞周质中存放硝酸盐还原酶;当此酶在细胞周质内存在时,不再对氧分子敏感,因此,好氧条件下也能发生反硝化作用。影响反硝化作用的主要因子也不再是氧,而是有机质和硝酸盐含量。当为反硝化作用提供底物,硝化和反硝化作用通常耦合发生。参与反硝化作用的微生物种类繁多,已发现有 80 多个属的细菌和部分古菌、真菌和放线菌均可能参与反硝化作用。
打破平衡之后
自然状态下上述三个过程是平衡的,但是如果人类向大气和土壤中释放了过量的氮素,这一平衡就会被打破。
这些微生物栖息在土壤里,原本与人类和平共处,相安无事;可是贪婪的人类首先打破了这种默契。为了应对日益增长的农畜产品需求,近百年来,农业生产中化肥使用量猛增,伴随着家用汽车的普及和石油燃料燃烧,氮元素向生态系统输入迅速增加。
大气氮沉降增加已成为重要的全球气候变化议题,沉降量不断增加所带来的生态问题日趋严重,大气氮沉降将增加铵根离子的硝化和硝酸根离子的淋失,进而引发森林土壤酸化、营养失衡、树木发育衰退、生物多样性减少和生产力降低。
科学家们对现有森林进行施氮肥模拟大气氮沉降,他们发现长期过量氮沉降能给土壤微生物带来负面影响;例如,改变微生物群体的种类、个别微生物种的数量和生活能力,同时也改变土壤微生物对植物枯枝落叶和根系的分解能力和速率。欧洲的科学家们研究发现森林氮饱和的临界负荷最小值为每年每公顷10千克氮,但目前欧洲中部地区大部分森林氮输入量为每年25到60千克,远远超出了森林对氮营养元素年均需求量。而在北美地区,某些森林生态系统的氮输入量每年每公顷达40千克。来自中国科学家的调查也不容乐观,每年降雨中所携带的硝酸根离子和沉降量与美国和日本等发达国家相接近,而降雨中的铵根离子浓度则更高,其沉降量也更大,分别是美国和日本的4倍和3.7倍。美国科学家在哈佛森林的松林林地的研究发现,微生物总量比未施氮林地减少了68%,阔叶林则减少了59%;同样,在维吉尼亚西部的2个氮饱和林地里亦发现了类似的现象,即微生物总量和真菌/细菌比率随着施氮水平增加而减少。另外,在英国北部的一个大草场样地,同样发现增施氮肥的草地土壤中真菌/细菌生物量比率显著低于未施氮的土壤。来自美国环保部国家健康与环境影响实验室的科学家们发现,与氮元素密切相关的微生物,特别是在高剂量氮肥处理的土壤中发现了大量的具有单加氧氨酶基因微生物,这是一类重要的硝化作用微生物,而在未增施氮肥的林地土壤中则没有发现;另外一种固氮细菌独有的基因在施氮肥土壤中没有发现,而在其它土壤中均有发现,这暗示了氮沉降增加已改变了固氮微生物类群。这些微生物群体的变化很可能会影响氮的轮回,进而影响我们的餐桌美食。(作者:郑俊强 姜萍)摘自中科院沈阳应用生态研究所网站,略删减